A l'écoute : Johannes-Passion de Johann Sebastian BACH (BWV 245, Nikolaus Harnoncourt et Arnold Schoenboerg Chor - Concentus Musicus Wien, 1995)
J'ai croisé la voix chaude et grave de Jean Claude Ameisen un jour de janvier 2010 sur France Inter. A l'instar de JSF (voir Réveil de la source) dont quelques mots m'avaient transpercé, les paroles de JC Ameisen m'ont littéralement soulevées ce jour là, et ouverts à des espaces qu'aujourd'hui encore je ne fais qu'explorer.
Jean Claude Ameisen (JCA) est Docteur en Immunologie et Professeur à l'Inserm et a écris en 1999
La Scuplture du Vivant paru au Seuil. Ce livre parle du concept de "mort cellulaire" en biologie et de ses liens avec l'embryogenèse et l'oncogenèse notamment, avant d'aborder les questions éthiques induites par ce concept fondamental et transversal peu à peu élaboré par de nombreux chercheurs.
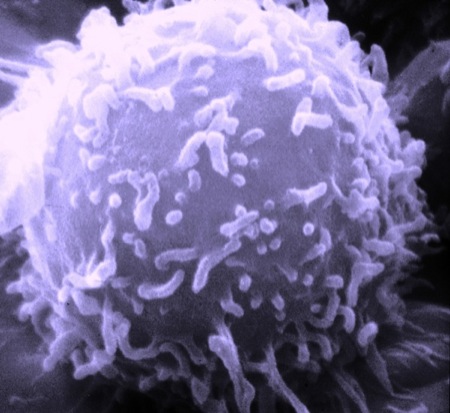
Dans ce premier billet sur la question, je souhaite présenter une image sur le système immunitaire qui m'a stupéfait. Mais d'abord le concept de mort cellulaire. Là encore, les concepts déployés ici sur la logique de l'antagonisme de S. Lupasco nous aident à prendre avec nous plus facilement cette complexité.
"Aujourd'hui, nous savons que toutes nos cellules possèdent le pouvoir, à tout moment, de s'auto-détruire en quelques heures. C'est à partir d'informations contenues dans leurs gènes que nos cellules fabriquent en permanence les "exécuteurs" capables de précipiter leur fin et les "protecteurs" capables de les neutraliser. Et la survie de chacune de nos cellules dépend, jour après jour, de sa capacité à percevoir dans l'environnement de notre corps les signaux émis par d'autres cellules, qui, seuls, lui permettent de réprimer le déclenchement de son autodestruction."
Ainsi JCA pose avec ces trois phrases emblématiques toute la beauté et la pureté du concept de "mort cellulaire" : il apparait ainsi un système complexe (et nous le verrons par la suite
infiniment complexe) où co-existent ensemble deux états antagonistes et opposés reliés indéfectiblement à leur environnement ouvert. Exécuteurs et protecteurs cellulaires co-existent ensemble dans un "équilibre" tendu à l'extrême, ouvert sur l'extérieur.
Le concept de vie est ici co-existant à celui de mort. Le concept de vie co-résulte de la "négation" du concept de mort comme ce dernier co-résulte de la négation du concept de vie. A ce jour, seule cette dernière partie de l'assertion nous parait "naturelle" et de bon sens. Et pourtant, la vie ne "triomphe" pas de la mort, elle en est humblement seulement sa co-partenaire.
Le concept de vie/mort dépend également de la présence continuelle de l'environnement, du même, comme du différent (non-même). Le concept de vie/mort ne se conçoit plus comme une entité solitaire mais indéfiniment solidaire. Une cellule ne peut vivre/mourir QUE si son environnement lui permet de réprimer son auto-destruction (et donc d'activer son auto-construction).
En langage lupascien, nous pourrions poser qu'au niveau cellulaire, la vie qui s'actualise (en réprimant les exécuteurs) potentialise en même temps la mort (par répression des protecteurs) et à la fois, la mort qui s'actualise (en activant les exécuteurs) potentialise en même temps la vie (par activation des protecteurs). Ce doublet d'antagonistes opposés s'ouvre et s'oppose à la fois dans le Tiers Inclus logique : l'interdépendance comme unique alternative à la vie/mort. Cette interdépendance absolue (au sens d'une contradiction entre vie et mort) fait émerger une co-dépendance relative (au sens d'une non-contradiction entre vie et mort) qui donne in fine à la vie/mort l'image "d'un sculpteur, au cœur du vivant, à l'œuvre, jour après jour, dans l'émergence de sa forme et de sa complexité"
"Rien n'a de sens en biologie, a écrit le biologiste
Dobzhansky, si ce n'est à la lumière de l'évolution." («
Nothing in biology makes sense except in the light of evolution. ») Ainsi pour "appréhender véritablement la "raison d'être" d'une propriété apparemment mystérieuse de nos cellules", il faut "partir à la recherche de ses origines."
Anne Fagot-Largeault dans ses cours au Collège de France en 2009 sur "l'Ontologie du devenir" souligne bien qu'en épistémologie des sciences biologiques et médicales, le philosophe comme le "praticien-chercheur" oublie fréquemment de considérer l'ontologie des concepts élaborés. Pour comprendre cela, JC Ameisen est parti à la recherche des origines du concept de mort cellulaire, dans les espèces les plus anciennes existant encore sur Terre : les bactéries, et a poursuivi sa quête tout au long de l'évolution du vivant jusqu'à l'homme.
L'ontologie retracée du concept de mort cellulaire aboutit ainsi à sa force et à consistance, à sa transversalité, sa transcendance. Nous reviendrons ultérieurement sur l'ensemble des retombées systémiques de ce concept dans diverses sciences...
Au chapitre 3 du livre vient l'analyse "de la mémoire et de l'identité" ou comment le concept de mort cellulaire vient ré-interpréter le système immunitaire. Dans ce système co-existent les trois notions d'identité, d'adaptation et de mémoire.
"La notion d'identité pré-suppose la capacité de distinguer entre des informations qui sont émises par notre corps (le soi) et des informations dont l'origine nous est étrangère (le non-soi). La notion de mémoire pré-suppose la capacité de distinguer entre des informations nouvelles, auxquelles nous n'avons jamais été confrontés auparavant, et des informations que nous avons déjà rencontrées. Se souvenir, c'est reconnaître; et reconnaître, c'est répondre différemment la deuxième fois de la première."
Cette dernière phrase résonne curieusement dans le contexte d'un traumatisme psychique : pour se souvenir, le sujet doit pouvoir répondre différemment par rapport à la première fois que l'évènement a surgi et a provoqué le "trauma". Ainsi, la réponse différente est la re-connaissance de cet évènement. Pour les théoriciens de l'EMDR par exemple, il existe une opposition entre principe de survie et principe de traitement de l'information : tant que l'information n'est pas "traitée" par la mémoire, au sens de sa signifiance pour le sujet, la survie seulement s'impose à lui (avec la cohorte symptomatologique qui y est liée : hyper-excitation somatique, syndrome de répétition, dissociation, évitement...etc).
Au niveau du corps humain, les notions d'identité et de mémoire ont une nature matérielle et concrète : "elles s'élaborent, s'inscrivent et s'incarnent dans des réseaux de cellules qui communiquent entre elles" ajoute JC Ameisen. Ce réseau de cellules forme un système de système et possède donc un état informationnel d'où émergent ces notions d'identité et de mémoire.
Dans le système immunitaire humain existent deux grandes familles cellulaires distinctes : les "sentinelles" et les lymphocytes, notamment les T, dont la "maturation" se produit dans le thymus pendant la vie embryonnaire.
Les cellules sentinelles circulent dans tout le corps et "fragmentent, découpent sans cesse une petite partie de toutes les protéines qu'elles contiennent". Ces fragments sont exposés à la surface de la cellule sur des présentoirs, tous identiques entre eux. L'information contenue dans ces présentoirs et dans les fragments de protéines récoltées dans tout le corps, en l'absence d'infection, constitue le "soi", l'identité immunitaire de la personne. Dès qu'une nouvelle infection a lieu, des fragments de protéines étrangères au corps sont, par le travail des "sentinelles", exposées sur leurs présentoirs. L'assemblage hétérogène composé du présentoir du "soi" et du fragment de protéine du "non-soi" est capturé par un des récepteurs d'un lymphocyte T qui se transforme en combattant pour lutter contre l'infection.
Comment le lymphocyte T "sait il" que cet assemblage est hétérogène, sachant qu'il n'a jamais rencontré auparavant aucune protéine issue d'un nouveau microbe ? En fait, il ne s'agit pas d'une reconnaissance suite à un apprentissage du non-soi (l'infinie variété des protéines étrangères à l'unicité de notre corps dont l'énumération exhaustive est impossible) mais bien d'un apprentissage de la nature de notre "identité" au moment où notre corps n'est qu'un embryon.
Les concepts de vie/mort vont se mettre à l'œuvre ici : la vie d'abord, dans l'exploration systématique du champ des possibles.
Le scientifique baigné dans la logique "classique" exclusive convoque ici le "hasard" pour expliquer l'élaboration d'un très grand nombre de combinaisons possibles d'un "jeu" combinatoire fini, en l'espèce la fabrication par l'embryon humain d'un nombre gigantesque de récepteurs
tous uniques mais tous différents de pré-lymphocytes T, qui comme "par hasard", vont par la suite définir parfaitement le "soi". La notion de "hasard", darwinienne, est pratique pour cacher le flou d'un mécanisme encore ignoré.
Mais il est possible aussi de voir ici un déploiement d'une systématique au sein d'une logique "inclusive", accompagné de manière concomitante d'une destruction d'une partie de ce qui a été préalablement déployé.
Je renvoie le lecteur tout d'abord à l'article
"Echelles, Nombres et Perception.." pour illustrer le principe d'une combinatoire systématique : combinatoire signifiant un calcul (addition et multiplication) et systématique signifiant que
tout est compté. JC Ameisen donne comme exemple un récepteur protidique composé de 4 protéines différentes codées par 600 gènes environ chacune. Si l'on admet une "fabrication" systématique de tous les récepteurs uniques possibles (différents d'au moins 1 protéine sur quatre), le possible devient vertigineux (environ 130x10^9 combinaisons possibles). Au sein d'une logique inclusive, ce possible définit un
potentiel. Ce potentiel n'existe en réalité que dans un tout petit organe au sein de l'embryon (le thymus) et pendant un laps de temps très court : cet organe est en fait comme un simulateur de vie ...et de mort potentielles.
Au sein du thymus de l'embryon sont "entreposés" tous les récepteurs des cellules sentinelles définissant le soi. Les récepteurs des pré-lymphocytes T vont durant trois jours circuler au sein de ces cellules et être "baignés" de ces informations, les cellules interagissant entre elles.
Comme nous l'avons vu plus haut, la survie du futur lymphocyte T dépend des interactions de son environnement. Ici s'opère l'étrange mais fascinante complexité de la construction de notre identité biologique. Il est possible, selon JC Ameisen, de simplifier les interactions complexes entre cellules selon deux groupes de réponses : les réponses qui vont permettrent la vie et celles qui vont donner la mort. Quantitativement, on constate que les réponses de mort seront beaucoup plus nombreuses que celles de vie et cela aboutit à la destruction par l'organisme lui même de près de 99% du potentiel de vie précédemment créé ! Les "raisons" de mort sont assez "binaires" : ainsi si les récepteurs du lymphocyte T interagissent trop fortement avec les récepteurs du soi, ils sont détruits (évitant ainsi les futures maladies auto-immunes); de même si les récepteurs n'interagissent pas du tout ou très faiblement, ils sont détruit également (évitant l'inutilité future du lymphocyte "tueur" !). En fait, seuls les récepteurs dont l'interaction est "modérée" vont permettre d'engendrer un signal "faible" de survie pour les lymphocytes T : ainsi la vie apparait ici comme la réponse "modérée" et "faible" : unique complexité située entre des réponses trop "franches", trop intenses, dans un sens comme dans l'autre.
Pendant trois jours, au sein du thymus, se joue ainsi l'actualisation du potentiel créé de manière systématique auparavant. Cette actualisation va garder environ 1% du potentiel et définir ainsi l'unicité singulière de l'embryon donc de l'individu en train de se former : singularité qui signera in fine sa "réponse" immunitaire.
Je vois pour ma part, ici, une complexité se mettre en place, au sein des relations (dynamiques) entre cellules de l'immunité. Je conjecture ici également un système de systèmes dont il est possible de comprendre l'ontologie si on s'empare de la logique lupascienne.
Posons que l'ensemble des interactions entre cellules sentinelles et pré- lymphocytes T dans le thymus embryonnaire définisse un ensemble de couples dont chacun est composé de deux informations : la combinaison du récepteur de la sentinelle et la combinaison du récepteur du lymphocyte T. Nommons pour simplifier notre propos chaque couple générique (S,T). Nous avons alors :
1) Si S est "trop proche" (interaction trop intense) de T, alors c'est la destruction de T
2) Si S est "trop éloigné" (interaction trop faible) de T, alors c'est la destruction de T
3) Si S n'est "ni trop proche ET ni trop éloigné" (interaction modérée) de T, alors c'est la survie de T.
Ce système axiomatique fait penser à une matrice ternaire où 1) serait "l'égalité" (axiome d'identité) (A est A), 2) serait la non égalité (axiome de non contradiction) (A n'est pas non-A) et 3) serait le tiers inclus (axiome du tiers inclus) (il existe un tiers entre A et non-A).
Pourquoi le tiers inclus ici et non l'axiome classique du tiers exclus ? Tout simplement car la réponse donne la "vie", c'est à dire actualise le potentiel existant en le gardant "visible", tandis que les autres réponses donnant la "mort" actualisent le potentiel existant en le faisant disparaître ! Le tiers inclus, ici dans cette systémogénèse immunitaire, définit la vie même tandis que les opposés contradictoires engendrent la mort. Il est bien entendu qu'ici, je parle des interactions entre cellules, donc de leurs relations dynamiques, c'est à dire in fine de systèmes informationnels et non des structures cellulaires au sens strict.
Que peut signifier ici un rapprochement entre le concept lupascien du tiers inclus et l'identité immunitaire du "soi" ? Je vois pour ma part l'irréductible contradictoire et l'inversion de sens (vu avec Van Gennep).
Selon JC Ameisein, la re-connaissance est une présentation à nouveau avec une réponse
différente. En effet, il écrit : "Aussi étrange que cela puisse paraître, le lymphocyte qui répond pour la première fois à un fragment d'une des protéines du virus de la grippe [...] n'y répond pas parce qu'il le "reconnait" [au sens de connaitre] : il y répond
au contraire pour la seule raison qu'il ne l'a jamais rencontré." Et cette réponse sera intense et cela déclenchera, pour la première fois, un signal intense qui engendrera la destruction de la protéine du "non-soi". Le "soi" est ici définit par des informations ( des combinaisons de protéines) "contradictoires" essentiellement, une réponse originelle modérée, singulière et rare, tandis que le "non-soi" se définit par des informations non contradictoires essentiellement, une réponse originelle très forte ou très faible, commune et courante (statistiquement du moins).
C'est bien l'inclusion originelle d'informations contradictoires, pendant la vie embryonnaire, qui permet de construire la complexité de notre système immunitaire.
Le concept de mort cellulaire apparaît ainsi très bien adapté à sa ré-interprétation au sein d'une métaphysique lupascienne de logique inclusive ou logique du tiers inclus. La complexité entrevue alors prend tout son sens et "se simplifie".
Ce concept de mort cellulaire, rend ainsi au concept de vie, toute sa beauté et sa complexité grâce à l'introduction du contradictoire au sein du couple (vie,mort). Immergé au sein d'une logique lupascienne, ce couple fait à son tour émerger peut-être
Le véritable concept de ce que nous nommons la "vie" : en réalité, son "tiers inclus", issu d'une tension irréductible et contradictoire entre vie et mort, et dont la manifestation "visible" reste si rare et si singulière !